

Genetic aspects of interspecies hybridization between sable and pine marten based on microsatellite loci data
* This article is part of a Special Issue from the Martes Working Group.

Abstract photo. Pine marten (Martes martes, left) photo by Alexander Gos’kov and sable (Martes zibellina, right) photo by Evgeniy Zakharov.
Abstract
Individuals with intermediate phenotype of the sable (Martes zibellina) and pine marten (Martes martes) are regularly observed in the sympatry zone of these species’ ranges. Presumably, these offspring are hybrids of sable and pine marten. We studied 246 animals, identified by morphological characters as 20 of pine martens and 226 of sables. We surveyed within areas focused on sympatry and identified neighboring territories based on genetic markers. In 6 out of 11 geographical localities of sable, we found individuals (N=10) with a significant portion of pine marten’s genotype. Among 20 samples of pine martens, we found one individual with a significant proportion of the sable’s genotype. Thus, accurate identification of sable and pine marten hybrids by morphological characters was practically impossible: in sympatric populations, some animals morphologically defined as sable or pine marten may be hybrids of these species.
Keywords: sable, pine marten, hybrid, kiduses, sympatry zone, microsatellite loci
Introduction
Sable (Martes zibellina) and pine marten (Martes martes) are two closely related species of the genus Martes, with genetic divergence about 1-2 Ma ago (Koepfli et al. 2008; Sato et al. 2012; Law et al. 2018). It has been assumed that sable originated in the Far East (Monakhov 1976), and the pine marten in Western Europe, possibly in the Alps (Grakov 1981). The modern range of the pine marten covers all of Europe, from the British Isles on the west to the Ural Mountains and the south of Western Siberia on the east. The sable range is mainly located in Asia, with the western edge of the northern Urals, and the eastern edge on the Far East, including the Kamchatka Peninsula and Okhotsk Sea islands. In the present, range of sympatry of the two species is located on the Northern Urals and on the southwestern part of Western Siberia (Fig. 1). It has been hypothesized that the range of the sable extended far to the west, reaching Scandinavia (Jurgenson 1956; Bakeyev et al. 2003; Monakhov 2022), and the pine marten in the east could have reached the basins of the Taz and Yenisei rivers (Monakhov 2022).
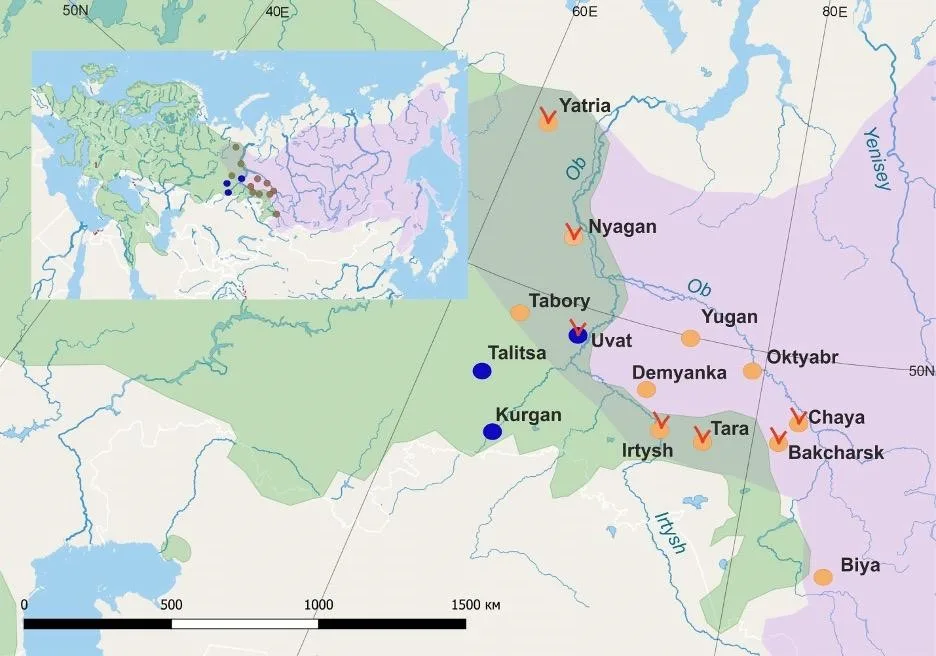
Figure 1. Sampled sites of pine martens (Martes martes, blue circles) and sables (Martes zibellina, yellow circles). Red marks indicate marten samples detected in this study as interspecies hybrids. The pine martens’ range are depicted in green color, sables’ range in purple color. Data on species ranges were based on Monakhov (2016) and Herrero et al. (2016) with modifications.
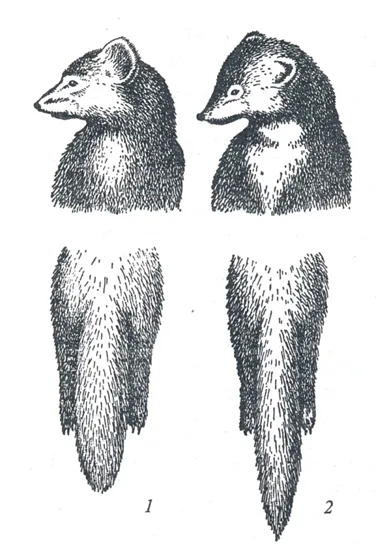
Despite the fact that sable and pine marten are well distinguished by external features (shape and coloration of the head, shape of the throat spot, tail length, fur of paws, Fig.2, Table 1) animals with intermediate phenotypic characteristics are often found in areas where both species are found together. Usually, such animals are identified as hybrids. The first references about the hybrid of sable and pine marten are in the late 18th century in the works of Pallas (1786), but a detailed description of the morphology and ecology of hybrids appears only in the first half of the 20th century (Manteifel 1934; Skalon et al. 1940, etc.). The first experiments on crossbreeding sable and pine marten in captivity were organized at the same time (Ponomarev 1946; Grakov 1974, 1981, 1993) when the first hybrid generations were obtained. The results of breeding experiments were inconsistent. In some cases, infertility of hybrids were described, and the phenomenon of heterosis was observed, when a hybrid male was much larger than parents (Ponomarev 1946). Other authors describe partial fertility of hybrids and intermediate characteristics of skull and fur size (Portnova 1941; Grakov 1974, 1981). Based on contradictory data on the description of wild hybrids, often referred to as kiduses in some journals (e.g., Monakhov and Uspenskaya 2013) , some have concluded the scale of hybridization between sable and pine marten was an overestimate, and hybrids of sables or pine martens would occur with a phenotype different from the majority of animals (Pavlinin 1963).
In previous studies, genetic markers, particularly microsatellites, have been used to identify hybrids (Pischulina 2013; Modorov et al. 2020). In this study, we attempt to assess the presence of hybrids in sympatric populations of sable and pine marten based on multiplex analysis of 11 microsatellite loci of nuclear DNA (Modorov et al. 2020).
Methods and Materials
This study was based on tissue samples (muscle or skin) of 216 hunted or trapped sables and 30 pine martens. None of the authors were involved in hunting, and no animal was killed with the aim of collecting samples for this work. All samples were obtained directly from licensed hunters. Species identification was made by hunters and based on morphological species-specific characteristics. Our samples included both new material (95 specimens) and individuals already analyzed in previous studies (Modorov et al. 2020; Ranyuk et al. 2021). Martens and sables were sampled at 14 sites of the study area (Fig. 1). DNA was extracted using the DNA-extran-2 Kit for Animals (Sintol, Russia). All the samples were genotyped at 11 microsatellite loci (Modorov et al. 2020). The sets of primers were chosen on the basis of already known microsatellite loci in the sable: Mzf51 and Mzf56 (Zhu et al. 2017); Martes americana and Gulo gulo Ma1, Ma3, Ma8, Ma15, and Ma19 (Davis & Strobeck 1998); Martes foina Mf8.7 and Mf8.8 (Basto et al. 2010); and Neogale vison, Mustela erminea Mvis72 and Mvi2243 (Fleming et al. 1999; Vincent et al., 2003). The selected microsatellite loci were located in different linkage groups (Modorov et al. 2020). The PCR was carried out as described by Modorov et al. (2020). PCR products were analyzed via automated capillary electrophoresis on an ABI 3130 (Applied Biosystems) genetic analyzer. Allele sizes were scored against an internal size standard S-550 Lyz (Gordiz) in GeneMapper v3.7 (Applied Biosystems).
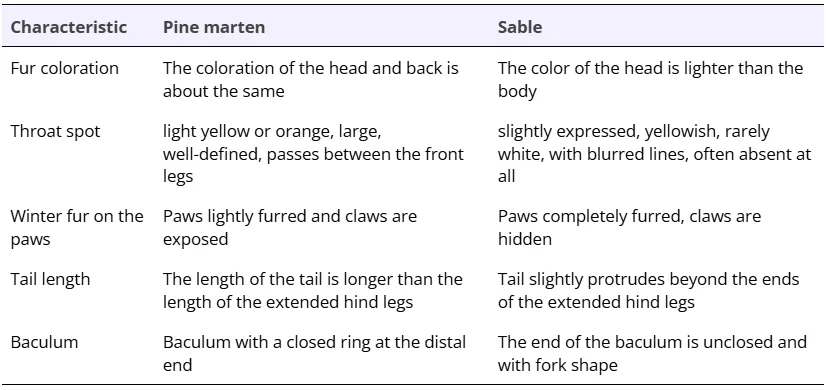
Table 1. Morphologic characteristics of sables and pine martens.
The observed (Na) and effective (Ne) number of alleles per locus and observed (Ho) and expected heterozygosity (He) (Nei, 1977) for the samples were estimated, and the analysis of molecular variance was performed using GenALEx 6.5 (Peakall & Smouse 2006, 2012).
We used STRUCTURE for accurate analysis of admixture-affected population structure. The program implements a model-based clustering method to infer population structure. We used Bayesian clustering realized in STRUCTURE v.2.3.4 (Pritchard et al. 2000) to infer gene pools (sets of alleles in a population) and for genotype assignment. The number of potential genetic clusters (K) was assessed via five individual runs for each K value ranging from 2 to 12 with a burn-in length of 10,000 and 100,000 iterations of each chain in accordance with the admixture model along with the assumption of correlated allele frequencies among groups (Falush et al. 2003). We examined the optimal number of clusters (K) by using two alternative approaches based on the change of mean log likelihoods of the data: LnP(D) of independent runs for K (Pritchard et al. 2000) and rate of change in the LnP(D), ΔK, between successive K values (Evanno et al. 2005). The outputs of 10 independent runs for optimal K values were integrated using CLUMPAK (Kopelman et al. 2015). The main pipeline function was used for the summation and graphical representation of the results obtained in STRUCTURE. For this research we identified individuals as “hybrid” if proportion of other species genotype (Q) at the K=2 was more than 0.1 (10%). This relatively low threshold value proved to be highly effective in previous studies of mammalian hybridization with microsatellites markers (van Wyk et al., 2017; Szatmári et al., 2021).
Results
For all 11 loci, no unidirectional deviations of the observed frequencies genotype from the expected frequencies according to the Hardy-Weinberg equilibrium were observed. Therefore, no loci were excluded for analyzing the genetic structure of the population.
The lowest values of the effective number of alleles per locus and heterozygosity were obtained for the pine marten sample (Table 2).
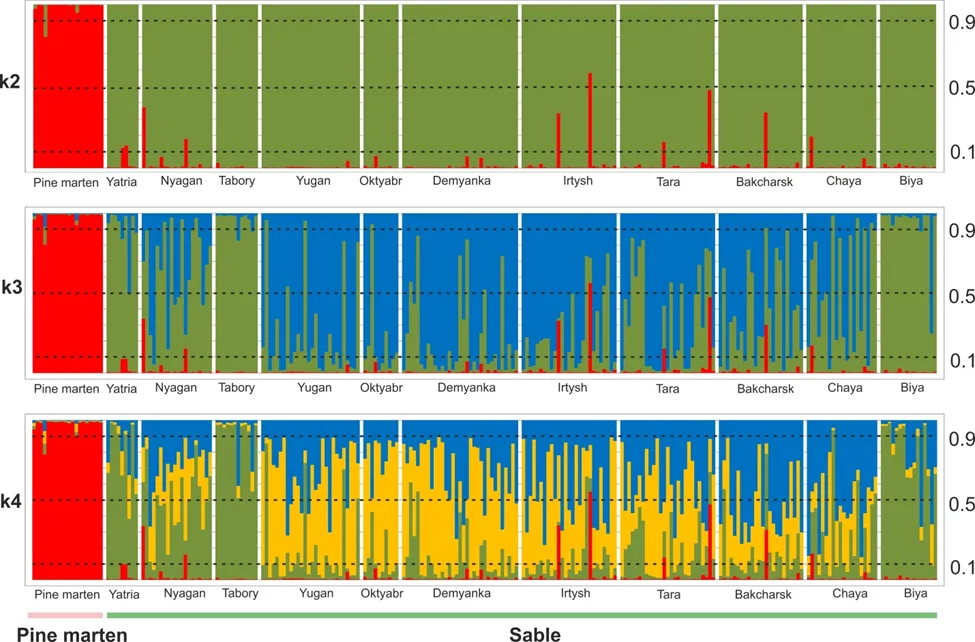
We provide Bayesian clustering results (K = 1–12) in Figure 3. The mean similarity score at K = 2–12 in the CLUMPAK main pipeline was high (0.89–0.99). Three genetic clusters were detected using Evanno’s method (the best K = 3). The highest value of LnP(D) detected for K = 4.
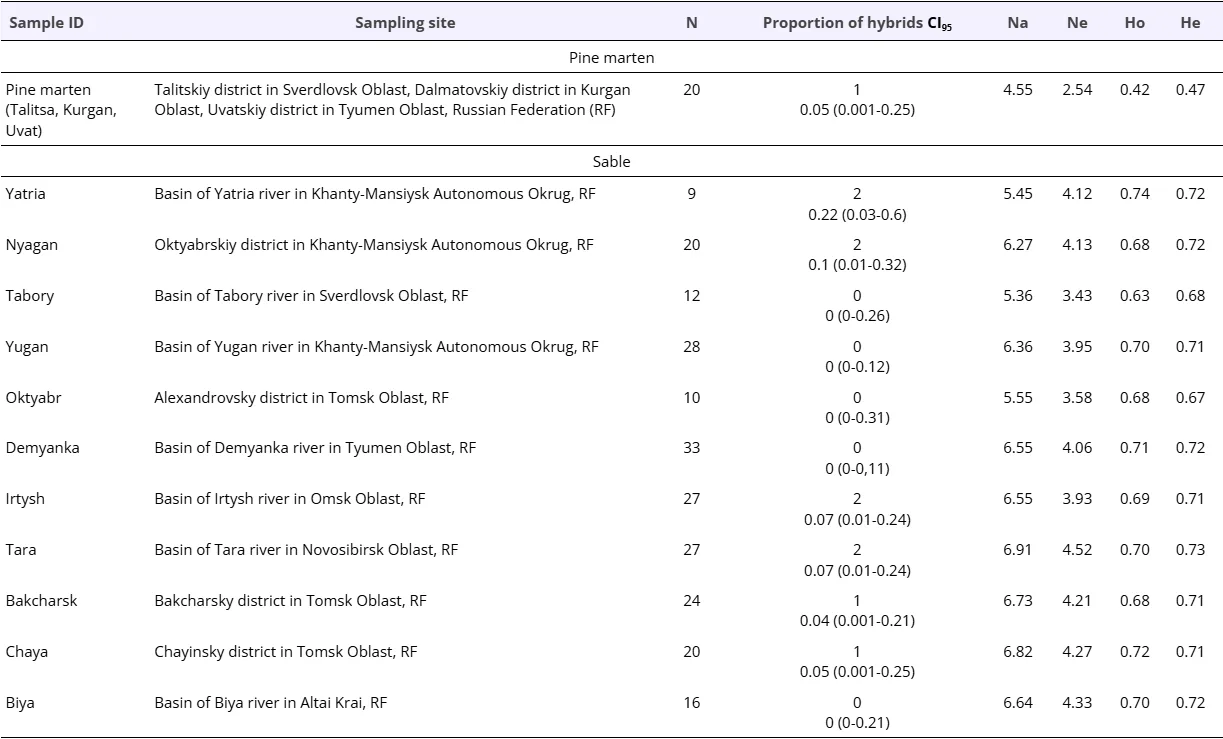
Table 2. Geographic locations of the populations analyzed in this study and a summary of genetic diversity metrics. Abbreviations: N, the number of individuals; CI95, confidence interval 95% (low bound - upper bound); Na, the mean number of alleles per locus; Ne, the effective number of alleles per locus; Ho, observed heterozygosity; He, expected heterozygosity. Three geographic locations of pine martens were pooled in one sample “pine marten” because of small number of individuals for Uvat (N=3) and Kurgan (N=3).
At K = 2, pine marten samples were separated from sable populations (Fig. 3). At the same time, individuals with more than 10% of the “pine marten” genotype were found in sable populations. Such individuals were found in samples Yatria, Nyagan, Irtysh, Tara, Bakcharsk and Chaya (Fig. 1 and 3). One individual with a high level of “sable” genotype was found in pine martens’ sample (Fig. 3). At K = 3 and K = 4 the population structure of the sable begins to differentiate where populations from the territory between Ob and Irtysh rivers are separated from others (Yugan, Oktyabr, Demyanka, Irtysh, Tara, Bakcharsk and Chaya).
Discussion
Considerable debate occurs regarding what defines a species (e.g., Dobzhansky 1935), but it has been argued that the biological definition of a species includes morphological, geographical, ecological, behavioral, and molecular information (Mayr 1996). Here, we explore molecular information to provide an assessment for two species – sable and pine martens.
Hybridization in a wild Mustelidae family has been recorded. There is genetic evidence of hybridization between two close related species of badgers Meles meles and M. leucurus in the contact zone (Kinoshita et al. 2018), natural hybrids of two polecat species, Mustela eversmanii and M. putorius were identified in the sympatric area (Szatmari et al. 2021). Interspecies hybridization was identified in two zones of contact between North American martens Martes americana and M. caurina (Colella et al. 2019).
The information about hybridization between the species of our study, Eurasian martens M. martes and M. zibellina dating back to the 18th century (Pallas 1786). There is even a special name for such hybrids – kidas or kidus. In the 21st century, not only genetic evidence of natural hybridization of these two close species appeared (Pischulina 2013, Modorov et al. 2020), but also the ability of such hybrids to produce fertile offspring were identified (Manakhov, 2022).
Sable and pine marten are typically thought to be well-distinguished by external features – head shape, paw’s fur, tail length, fur coloration and properties (Fig. 2, Table 1). Craniological, osteological and odontological characters have a large zone of overlapping for sables and pine martens and generally are not considered suitable for individual species identification (Gasilin, Kosintsev 2013; Gimranov, Kosintsev 2015; Ranyuk, Monakhov 2016). Genetic markers, particularly microsatellite loci and mtDNA, can be used as additional evidence to differentiate species (Rozhnov et al. 2010; Modorov et al. 2020).
The multiplex of 11 microsatellite loci used in this study was quite successful in separating species from each other (Fig. 3). Pine marten has a low genetic variability compared to sable (Table 2). Similar results have been obtained previously for other genetic markers (Manakhov 2022) and morphological traits (Monakhov 2021). This could be due to differences in species ecology where the pine marten may be adapted to a temperate climate, whereas the sable appears more adapted to cold and extreme climates. Another difference could be that sables were described with specific intraspecies genetic structure in the species’ range (Ranyuk et al. 2021), whereas pine marten from the whole East European Plain formed one genetic group (Manakhov 2022). Massive translocations carried out in the middle of the 20th century, when sables were moved from the Baikal area to Western and Eastern Siberia (Pavlov et al. 1973) could additionally increase the genetic diversity of sables.
Population structure of the sable was also revealed in our study when according to Bayesian clustering the samples from West Siberian Plain formed the one genetic cluster at K=3 and K=4 (Fig. 3). Presumably rivers Ob and Irtysh play the role of some landscape barriers for dispersal ability of the animals. Samples from Yatria, Tabory and Biya formed an opposite genetic cluster (Fig. 3). Sable showed a high level of the genetic variability in common and it can be related to high migration activity of the species (Bakeyev et al., 1980; Chernikin, 2006). Previous studies of the sable’s genetic structure showed positive autocorrelation occurs in the Mantel correlogram in 300–600 km distance class (Ranyuk et al. 2021). Samples Yatria and Tabory from the western bank of Ob River and population from the basin of Biya River were separated from each other about 1500 km and their presence in the common genetic cluster cannot be explained by migrations only. All these samples are from the autochthonous sable’s populations so translocations could not influence the genetic structure of these samples. Intraspecies structure of the sable is a very complicated and interesting problem and to bring light to this question we need more geographic samples and additional genetic markers.
Here, we compared animals identified to species by experienced hunters based on morphological characteristics as pine martens or sables. According to our genetic analysis in 6 geographic regions, samples of ‘sable’ individuals also exhibited a significant level of the pine marten genotype, which may suggest that these are interspecific hybrids. Our sampled animals did not differ in morphology from the sables harvested on the same territory. Among the pine martens sampled, an individual with a high part of the sable genotype also was found. Our data confirm the results of a previous genetic study of sables and pine martens from the Northern Urals where it was difficult to identify specific individuals in the sympatry zone of the pine marten and sable by physical characteristics, since the individuals of parental species deviated in morphological traits (Pischulina 2013). Some genetic hybrids, on the contrary, did not differ from one of the parental forms. Genetic methods identified up to 20% of individuals in the sympatric population of the Northern Urals as hybrids, but the results of phenotypic and genetic identification of hybrids did not coincide in 37% of cases (Pischulina 2013). In our study we included individuals with morphological characters of “pure” species and interspecies hybrids were found in half of geographic samples with proportion from 4% (Bakcharsk) to 22% (Yatria). Thus, accurate identification of sable and pine marten hybrids by morphological characters is practically impossible: in sympatric populations, some animals morphologically defined as sable or pine marten may be hybrids of these species, while some individuals morphologically identified as hybrids may be representatives of one of the species.
In the IUCN Red List, pine marten and sable have been assigned to the ‘Least Concern’ category (LC) (Herrero et al., 2016; Monakhov, 2016). However, these two Martes species are hunted, and there is always the threat of over-harvesting of certain populations. The sable’s and marten’s populations from the sympatry zone are unique and following the strategy of biodiversity conservation at the intraspecific level, special attention should be paid to the martens and sables in Ural, Western Siberia and Altai Krai.
Acknowledgments
The authors have no acknowledgements.
Author Contributions
Maryana N. Ranyuk: laboratory work, data analysis, manuscript preparation
Makar V. Modorov: laboratory work, data analysis, manuscript preparation
Vladimir G. Monakhov: collection of material, manuscript preparation
Data Availability
Data and R code will be made available upon request.
Transparent Peer Review
Results from the Transparent Peer Review can be found here.
Recommended Citation
Ranyuk, M.N., Modorov, M.V., & Monakhov, V.G. (2025). Genetic aspects of interspecies hybridization between sable and pine marten based on microsatellite loci data. Stacks Journal: 25005. https://doi.org/10.60102/stacks-25005
References
Bakeyev NN, Monakhov GI, Sinitsyn AA. 2003. The sable. All-Russian Institute of Hunting and Fur Farming Press, Vyatka (in Russian)
Bakeyev NN, Chernikin EM, Shilyaeva LM. 1980. Carnivore, in: Kucheruk V.V. (Ed.), Results of marking on mammals. Nauka Publishing House, Moscow, Soviet Union, pp 77–94 (in Russian)
Basto MP, Rodrigues M, Santos-Reis M, Bruford MW, Fernandes CA. 2010. Isolation and characterization of13 tetranucleotide microsatellite loci in the stone marten (Martes foina). Conserv Genet Resour 2: 317–319 https://doi.org/10.1007/s12686-010-9217-2
Chernikin EM. 2006. Ecology of Sable (Martes zibellina) in Barguzin Reserve. Buryat State University Press, Ulan-Ude, Russia (In Russian)
Colella JP, Wilson RE, Talbot SL, et al. 2019. Implications of introgression for wildlife translocations: the case of North American martens. Conservation Genetics 20:153–166 https://doi.org/10.1007/s10592-018-1120-5
Davis CS, Strobeck C. 1998. Isolation, variability, and cross-species amplification of polymorphic microsatellite loci in the family Mustelidae. Mol Ecol 7(12): 1776–1778 https://doi.org/10.1046/j.1365-294x.1998.00515.x
Dobzhansky T. 1935. A critique of the species concept in biology. Philosophy of Science 2:344-355
Evanno G, Regnaut S, Goudet J. 2005. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14(8): 2611–2620 https://doi.org/10.1111/j.1365-294X.2005.02553.x
Falush D, Stephens M, Pritchard JK. 2003. Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies. Genetics 164(4): 1567–1587
Fleming MA, Ostrander EA, Cook JA. 1999. Microsatellite markers for American mink (Mustela vison) and ermine (Mustela erminea). Mol Ecol 8(8): 1352–1354 https://doi.org/10.1046/j.1365-294X.1999.00701_2.x
Gasilin VV, Kosintsev PA. 2003. Diagnosis of species of the subgenus Martes str. (Carnivora, Mustelidae) by metric features of the mandible. Zoologocheskiy Zhurnal 92(2): 221–230 (in Russian)
Gimranov DO, Kosintsev PA. 2015. Differentiation of three Martes species (M. martes, M. zibellina, M. foina) by tooth morphotypes. Comptes Rendus Palevol 14: 647-656
Grakov NN. 1974. On the species independence of the pine marten and sable. Proceedings of All-Union Research Institute of Hunting 46: 11–14 (in Russian)
Grakov NN. 1981. The pine marten. Nauka Publishing House, Moscow, Soviet Union (in Russian)
Grakov NN. 1993. Pine Marten and its harvest in Russia. Lutreola 2: 7–13
Herrero J, Kranz A, Skumatov D, Abramov AV, Maran T, Monakhov VG. 2016. Martes martes. The IUCN Red List of Threatened Species 2016:e.T12848A45199169
Jurgenson PB. 1956. Essays on the comparative study of sable and martens. Materials on the results of mammal studies in State Reserves. Moscow, pp33–71 (in Russian)
Kinoshita E, Abramov AV, Soloviev VA, et al. 2019. Hybridization between the European and Asian badgers (Meles, Carnivora) in the Volga-Kama region, revealed by analyses of maternally, paternally and biparentally inherited genes. Mamm Biol 94: 140–148 https://doi.org/10.1016/j.mambio.2018.05.003
Koepfli K-P, Deere KA, Slater GJ, et al. 2008. Multigene phylogeny of the Mustelidae: Resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biol 6:10 https://doi.org/10.1186/1741-7007-6-10
Kopelman, NM, Mayzel J, Jakobsson M, Rosenberg NA, & Mayrose I. 2015. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Molecular Ecology Resources 15(5): 1179–1191 https://doi.org/10.1111/1755-0998.12387
Kuznetsov BA. 1975. Field guide of the vertebrates in the fauna of USSR. Mammals. Moscow, Prosveschenie (In Russian)
Law CJ, Slater GJ, Mehta RS. 2018. Lineage diversity and size disparity in Musteloidea: testing patterns of adaptive radiation using molecular and fossil-based methods. Syst Biol 67: 127–144 https://doi.org/10.1093/sysbio/syx047
Manakhov AD. 2022. Genomic Analysis of Mustelidae Family Representatives. Dissertation, N.I. Vavilov Institute of General Genetics, Russian Academy of Sciences. Moscow (in Russian)
Manteifel PA. 1934. The sable. KOIZ, Moscow–Leningrad, Soviet Union (in Russian)
Mayr E. 1996. What Is a Species, and What Is Not? Philosophy of Science 63(2): 262–277
Modorov M, Monakhov V, Mikryukov V, et al. 2020. Microsatellite multiplex assay for sable (Martes zibellina) and pine marten (Martes martes). Mamm Res 65: 855–862 https://doi.org/10.1007/s13364-020-00529-4
Monakhov GI. 1976. Geographic variability and taxonomic structure of Sable of USSR fauna. Scientific Bulletin of the Institute of Animal Origin and Furs (Kirov, Soviet Union) 26: 54–86 (in Russian)
Monakhov VG. 2011. Martes zibellina (Carnivora: Mustelidae). Mamm Species 43(1): 75–86 https://doi.org/10.1644/876.1.
Monakhov VG. 2016. Martes zibellina. The IUCN RedList of Threatened Species 2016:e.T41652A45213477 https://doi.org/10.2305/IUCN.UK.2016-1.RLTS.T41652A45213477.en
Monakhov V. 2021. Tracing size distribution patterns in two Martes species across Eurasia: which populations should be protected? Biodivers Conserv 30: 37–53 https://doi.org/10.1007/s10531-020-02072-9
Monakhov V. 2022. Martes martes (Carnivora: Mustelidae). Mamm Species 54: 1–22 https://doi.org/10.1093/mspecies/seac007
Monakhov VG, Uspenskaya OD. 2013. On the Morphological Distinctness of the Hybrid between the Sable and Pine Marten. Doklady Biological Sciences 448: 732–736 https://doi.org/10.1134/S0012496613010171
Nei M. 1977. F-statistics and analysis of gene diversity in subdivided populations. Annals of Human Genetics 41: 225–233 https://doi.org/10.1111/j.1469-1809.1977.tb01918.x
Pallas PS. 1786. Traveling through different provinces of the Russian Empire. Part II. Saint-Petersburg, Imperial Academy of Sciences (in Russian)
Pavlinin VN. 1963. Tobolsk Sable. Ural Branch of Academy of Sciences of USSR, Sverdlovsk, Soviet Union (in Russian)
Pavlov MP, Korsakova IB, Timofeev VV, et al. 1973. Acclimatization of game mammals in the Soviet Union. Part 1. pp 51–105 (in Russian).
Peakall R, Smouse PE. 2006. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6(1): 288–295 https://doi.org/10.1111/j.1471-8286.2005.01155.x
Peakall R, & Smouse PE. 2012. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research – An update. Bioinformatics 28: 2537–2539 https://doi.org/10.1093/bioinformatics/bts460
Pishchulina SL. 2013. Relationships between pine marten and sable in sympatry zone: genetic aspect. Dissertation, Institute of ecology and evolution of the Russian Academy of Sciences (in Russian)
Ponomarev AL. 1946. Kidus. Bulletin of the Moscow Society of Nature Researchers 51(4-5): 79–83 (in Russian)
Portnova AT. 1941. Breeding of kidus. Rabbit breeding and fur farming 6: 22–23 (in Russian)
Pritchard JK, Stephens M, Donnelly P. 2000. Inference of population structure using multilocus genotype data. Genetics 155(2): 945–959
Ranyuk MN, Monakhov VG. 2016. A comparative analysis of craniological variability of Eurasian (sable and pine marten) and North American (American marten) species of the Martes genus. Biology Bulletin 43(8): 893–903
Ranyuk M, Modorov M, Monakhov V, Devyatkin G. 2021. Genetic differentiation of autochthonous sable populations in Western and Eastern Siberia. Journal of Zoological Systematics and Evolutionary Research 59: 2539–2552 https://doi.org/10.1111/jzs.12565
Rozhnov VV, Meschersky IG, Pishchulina SL, Simakin LV. 2010. Genetic analysis of sable (Martes zibellina) and pine marten (M. martes) populations in sympatric part of distribution area in the northern Urals. Russ J Genet 46(4): 488–492 https://doi.org/10.1134/S1022795410040150
Sato JJ, Wolsan M, Prevosti FJ, et al. 2012. Evolutionary and biogeographic history of weasel-like carnivorans (Musteloidea). Mol Phylogenet Evol 63: 745–757 https://doi.org/10.1016/j.ympev.2012.02.025
Skalon VN, Rayevskiy VV, Zhbanov YeS. 1940. Current distribution and interactions of sable and pine marten in the north-eastern Urals. Scientific and methodological notes of the Committee on Reserves 7: 157–165 (in Russian)
Szatmári L, Cserkész T, Laczkó L, et al. 2021. A comparison of microsatellites and genome-wide SNPs for the detection of admixture brings the first molecular evidence for hybridization between Mustela eversmanii and M. putorius (Mustelidae, Carnivora). Evolutionary Applications 14:2286–2304
Van Wyk AM, Dalton DL, Hoban S, Bruford MW, Russo IRM, Birss C, Grobler P, van Vuuren BJ, & Kotzé A. 2017. Quantitative evaluation of hybridization and the impact on biodiversity conservation. Ecology and Evolution 7: 320–330 https://doi.org/10.1002/ece3.2595
Vincent IR, Farid A, Otieno CJ. 2003. Variability of thirteen microsatellite markers in american mink (Mustela vison). Can J Anim Sci 83(3): 597–599 https://doi.org/10.4141/A03-001
Zhu WC, Wei QG, Xue SY, Zhang HX, Lv TS, Zhang HH. 2017. Isolation and characterization of microsatellite markers for the sable, Martes zibellina (Mammalia: Mustelidae). Pak J Zool 49(5): 1909– 1912 https://doi.org/10.4238/gmr.15048
Open Access
Peer-Reviewed
Creative Commons
Accepted: 17 April 2025
Published: 23 May 2025
Funding Information: The study was carried out with the financial support of the Russian Foundation for Basic Research (project 122021000084-4).
Conflicts of Interest: The authors declare no conflicts of interest.



